
Picture – The idea of Viera da Silva’s art as a kind of code to be decoded comes across most clearly in The Chess Game – “O jogo de Xadrez” (above, Oil on Canvas, 1943). […] The checkered pattern of the chessboard extends beyond the table not only to the players themselves but also to the very landscape itself […] … Vieira da Silva would have loved The Matrix films […] (more).
Last night I decided to do something new. To play and broadcast live on Twitter, two chess games, blindfold. A 1st one with white pieces, another playing black. For that, I have chosen Chess Titans (link) has my contender, a computer program most people can also access and try out over their PC’s. Chess Titans is a computer chess game developed by Oberon Games and vastly included in Windows Vista and Windows 7. While broadcasting the game live, I added some of my thoughts while playing both games. Even if in brief, that was what I was feeling at the moment: what I was planning, and in what adversary menaces I mostly decided to spent my time.
For those reasons, what follows are those on-the-fly live comments, uncut, made at each moment, while I was thinking. No extra analysis is included here today. It will be more interesting for those who will read me on the future, I guess. This could give a precise idea what happened each time I have made a move, how I react it to some computer moves, and how some of my errors happened as you will see. How my mind went in one direction, or several, depending on the position. Those comments are highlighted by brackets () below, and were twitted live as they arrived to me. Besides, two subsequent comment brackets do not mean two subsequent twitter live chess thinking comments. Sometimes, several minutes have passed between those different thoughts.
As a final note, Chess Titans played each move in around 15-35 seconds, and in difficult positions, rarely, up to 2-3 minutes (I have chosen to play against the maximum level, 10). Playing blindfold, I have spent around 3-4 minutes for regular moves, like exchanging pieces, tweeting, etc, and mostly around 10-15 minutes for some positions, in quite difficult combinatorial patterns. First game playing white, endured 1h and a half (lost it) ,while the second almost 4h and 30 minutes within 58 moves. Here they are:
Game (1) Vitorino Ramos vs. Chess Titans level max.=10 [Sicilian] (LIVE on Twitter 23:00 GMT – 00:24 GMT, Dec. 20, 2012) Duration: 1h 24m.
1. e4, c5 2. c3, Nf6 3. Qc2, e5 4. Ne2, d5 5. exd5, Qxd5 (hmm … 6. d4 or 6.Ng3) 6. d4, Nc6 (7. c4 8. d5 but feeling problems later with his Nb4, Qa4+, Bd7!) 7. dxe5, Nxe5 8. Nf4, Qd7 9. Na3 (for 10. Bb5!), 9. …, Qe7 (was expecting 9. … a6) (10. Be3 seems too bad. Maybe 10. Be2 or Qe2. Or the line 10. Bb5+, Bd7, BxB, Nexd7+, Be3, Ng4 hmm … then Nd5!! ok … 10. Bb5+) 10. Bb5+, Bd7 11. 0-0, (better than BxB+ I guess cause of a future Ng4 by him), 11. …, g5

Chess diagram – crucial position after his 11. …, g5 move. White (me) to play.
(too risky maybe 12. Re1, gxN, Bxf4, Nf6-g4, f3 difficult for me to compute the rest) (12. Re1, gxN, Bxf4, Nf6-g4, f3, … hmm … Nexf3+ ?!!!)
(how about h3; 12. Re1, gxN, Bxf4, Nf6-g4, h3) (hmm???? 12. Re1, gxN, Bxf4, Nf6-g4, h3, Nxf2, Kxf2, Neg4+ ~ hmm) (we also have intermediate variants like, Bxb5, Nxb5, Q moves and gains one tempo by attacking the Knight on b5) (ok, no prob, here I go. This will be bloody …)
12. Re1 12. …, Nf3+ (Oooohhh NO!!!! damn, calculated this more ahead, not now. So stupid) 13. gxf3, Qxe1+ 14. Kg2, gxf4 (now he has Rg8++) (Bxf4 for Rg8+, Bg3 he has QxRa1, bad, bad) 15. Bxd7+, Nxd7 (h3 is an escape now for my King) (16. Rb1, Rg8+, Kh3, Qf1+, Kh4, Be7+ and I think I’m lost) (k, let’s sacrifice the Rook in a1) 16. Bxf4, Qxa1 (at least I have some counter-game now) 17. Qe4+, Be7 (Bd6 will not work due to Rg8+ followed by Qf1+ I guess…, damn, should have played 17. Qe2+!!) (Nb5 menacing Nc7+ or Nd6+ does not work either. Follows Rg8+, Kh3, Qf1+ and Q takes Nb5) (and for Qxb7 he has the robust Rb8 answer) (…. k, the end. Give up. Chess Titans level 10 won 1st game – 2nd game follows) 0-1
After two big blunders on the first game above (the bad 12. Re1 instead of a normal 12. Nd3 – check 1st diagram above -, and 17. Qe4+ instead of 17. Qe2+, since controlling f1 was crucial) the second game did not started well also. After 6 moves I was already losing 1 pawn. Yet, still did manage to open the game and get the initiative a few moves later (around 14. …, Re8+). I feel OK with open and highly combinatorial games as these (normally it’s when I play better), but I forgot one simple fact: I was playing blindfold. Four and an half hours later I guess I’m happy to have managed to drawn a quite interesting and complex game, playing black pieces. What a long and stressful headache. Here:
Game (2) Chess Titans level max.=10 vs. Vitorino Ramos [English opening] (LIVE on Twitter 00:45 GMT – 05:12 GMT, Dec. 20, 2012) Duration: 4h 27m.
1. c4, e5 (English) 2. Nc3, f5 3. g3, c6 4. e4, fxe4 5. Qh5+ (that 4. …, fxe4 was too bad from me. Childish error. Did not see the typical Qh5+ trap, g6, Qxe5+ followed by QxRh8. It should have been 4. …, d6) 5, …, Ke7 6. Qxe5+, Kf7 7. Be2, Qf6 8. Qxe4, Bc5 9. Nf3, Nh6 10. d4 (hmm prepares Ng5+ ??!) 10. …, Bb4 11. Bg5, Qf5 12. Qxf5+, Nxf5 13. Bd3, d6 14. a3, Re8+ 15. Be2, Bxc3+ 16. bxc3, h6 17. Bd2, g5 18. h4, g4 19. Nh2, h5 20. Bf4, b5 21. cxb5, cxb5 22. f3, Bb7 23. Rf1, gxf3 24. Nxf3, Nd7 25. Kd1, a6 26. Ng1, Kg6 27. Re1, Rac8 28. a4 (hmm … Bxh5+ is dangerous if I move the rock in column c, like 28. …, Rxc3), 28. …, Nf6 29. axb5, axb5, 30. Ra7, Bc6 (did calculate Ba8 and Bd5 but hmm, I need d5 for my knight. His bishop on f4 must die) 31. Bd3, Nd5 32. Ne2

Chess diagram – position after his 32. Ne2 move. Black (me) to play. I’m 1 pawn down but with the initiative.
(I can’t take on c3 right? Nxc3, Nxc3, Bf3+, and then he goes back with Knight to e2, gee…) (hard position to mentally calculate) (32. …, b4 ?????) (damn, let me simplify all this…) 32. …, Ra8 33. Rxa8, Rxa8 34. Bxd6 (geee, that 31. Bd3 was so well played) 34. …, Ra1+ (will try to drawn him with successive pressure and checks, I guess) 35. Kd2, Ra2+ 36. Kc1 (yep, he prepares to play Bb1, I guess) 36. …, Nde3 37. Nf4+, Kf7 38. Nxh5, Ra1+ 39. Bb1 (only move for him. If not I change the rocks in e1 with time and then his bishop on d6) 39. …, Be4

Chess diagram – position after my 39. …, Be4 move, pinning b2. White (computer) to play. I’m now 3 pawns down.
(Pinning. Guess this would end with 2 knights and 1 pawn against 1 knight and 4 pawns!!) 40. Kb2, Rxb1+ 41. Rxb1, Bxb1 42. Kxb1, Nxd6 43. Nf4 …
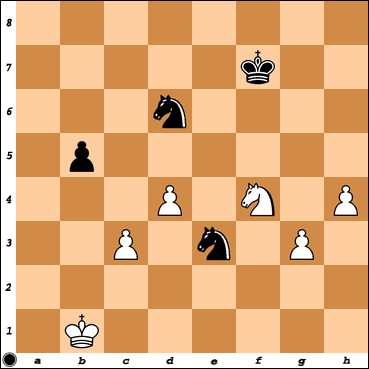
Chess diagram – position after his 43. Nf4 move. Black to play. Now I must stop two different white pawn clusters, on each side. Hard final.
(must be careful, now) (I guess I will do the obvious) (hmm, does not work, 43 …. Ne4 44. Ne2!) (wait, then King on f6, f5, g4 pressing g3) (k, here I go) 43. …, Ne4 44. Ne2, (now, I must think of my pawn on b5, hmm) (he has Ka2, a3 etc) (I have Nc4-d6, hope this helps, … here I go) 44. …, Kf6 45. Kb2, Kf5 46. h5 (?????!!!) 46. …, Kg5 47. h6 (?? He wants my King outside the centre, is that it? … I must take it) 47. …, Kxh6 48. Kb3 (yep, now I have problems on the other side) 48. …, Nd6 49. Kb4 (now my aim will be to arrive on f3 with my King) 49. …, Kg5 50. Kc5, Nec4 (freezing everything!) 51. d5 (hmm, I get it, he wants to reach Kd4 and Kd3. Anyway, I will go for the one in g3) 51. …, Kg4 52. Kc6
(what?????? he is just waiting) (hmm … wait, makes some sense. If 52…, Kf3 then 53. Nd4+, Kxg3 54. Nxb5, Nxb5 55. Kxb5 and I would have 1 knight against 2 pawns and my King far away) (hmm, hard call) (52…, Kf3 or not 52…, Kf3 ??!!!) (Kf3 followed by Ke3 and Kd3 etc does not work also, I think) (… hmm, wait, it might if he does not go Kc5, Kd4. If he goes I will the other way around by Kf4, Ke5)
52. …, Kf3 53. Nd4+, Kxg3 54. Kc5, Kf4 55. Nxb5, Ke5 (and it’s a drawn, I guess) 56. Kb4, Nxb5 57. Kxc4 57. …, Nxc3 ( if he goes 58. d6 then 58. …, Nd5! 59. d7, Nb6+ followed by Nxd7!!) 58. Kxc3, Kxd5 ½–½ (uuuufff, managing to draw blindfold, is a good result I guess :)
One of my conclusions: never play blindfold again in a open and highly combinatorial position, namely when you have a pair of knights. That, could make you dizzy and sick. Another (among, many others): never live tweet chess again. You will loose a lot of dumb followers (which turns-out to be healthy) and simultaneously attract all kinds of weirdos, and guru-like spam on-line marketeers. Vieira da Silva was right. It extends beyond the table. Like lake ripples when a stone is thrown.























Recent Comments